

Discover the significance of nutrition analysis in our health-conscious world through our comprehensive guide. We explore its role in aiding informed dietary decisions, assisting food manufacturers, and enforcing labeling laws. Learn about the FDA's pivotal role in shaping nutrition analysis practices. We discuss various analysis methods, including food testing and software-based calculations. Empower yourself to make healthier choices and support the development of nutritious products. Join us in unraveling the secrets of nutrition analysis for a healthier tomorrow.

Water quality is important for public health, and total coliform testing plays a role in ensuring the safety of our drinking water. We will explore what total coliform testing entails, its significance, interpretation of results, and when it's necessary.
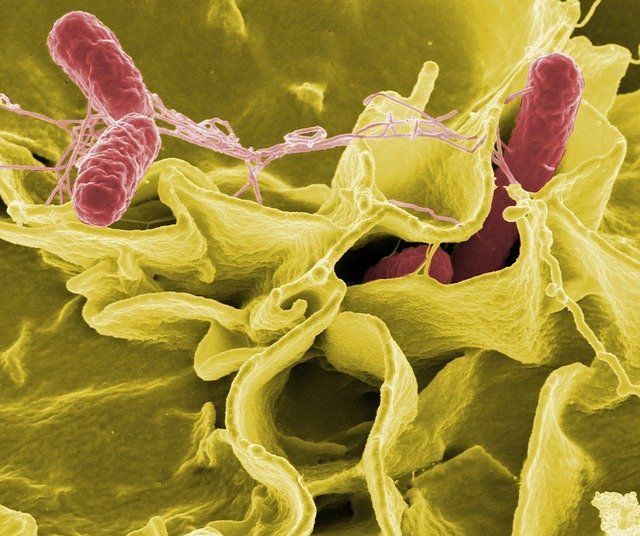
The significance of Bacillus cereus testing in ensuring food safety! Found in soil, dust, and raw foods, this bacterium poses a significant concern due to its potential to cause foodborne illness. Discover how food and environmental laboratories conduct rigorous testing to assess the safety and quality of food products, crucial for industries involved in food production and healthcare settings. Explore the methods, including microbiological culture and molecular techniques, employed for accurate detection. Join us in understanding how Bacillus cereus testing plays a pivotal role in upholding food safety standards, ensuring that consumers receive safe and high-quality food products.

From "best before" dates to expiration labels, shelf-life testing ensures the freshness and quality of our food and beverages. Delve into the world of quality control measures like microbiological, real-time, chemical, comparative, and accelerated shelf-life studies conducted in food safety laboratories. Learn how factors such as composition, storage conditions, and packaging influence product durability. Join us in appreciating the meticulous science that guarantees the safety and nutritional value of the products we enjoy every day!

Discover why this microbiological testing method is crucial for assessing microbial contamination in process water used across industries like pharmaceuticals and food production. Learn about the HPC test process, from sample collection to interpreting results, and its role in ensuring quality assurance, health and safety, and regulatory compliance.

Food is an essential part of our daily lives, providing sustenance and enjoyment. However, the safety of the food we consume can sometimes be compromised by the presence of pathogenic microorganisms. These unseen threats, invisible to the naked eye, can cause foodborne illnesses with potentially severe consequences. In this blog, we delve into the world of pathogenic microorganisms in food, their sources, impact on human health, and the crucial role of food safety measures in preventing their spread.
From understanding what air quality entails to exploring the factors contributing to pollutants, we'll unravel the importance of air quality testing and steps you can take to maintain clean air. Discover how pollutants from natural and human activities impact air quality and why frequent testing is crucial for health protection, awareness, and evaluating pollution control efforts. Ensure the air you breathe is clean and safe with Sure BioChem Laboratory's air quality analysis services. Take action to minimize pollutant-emitting products, ensure proper ventilation, and stay updated on-air quality conditions in your area.

Cleanrooms are a fascinating and critical aspect of modern industries that require precise and contamination free environments. These controlled spaces play a pivotal role in various sectors, ranging from pharmaceutical and biotechnology to electronics and aerospace.
In this post, we will delve into the world of cleanrooms in pharmaceutical and biotechnology, exploiting their definition, importance, and the technologies that ensure the highest standards of cleanliness.

Air monitoring is an important and routine aspect of maintaining a healthy cleanroom space. Take a close look at what airborne environmental monitoring systems are, how they work, and why they are important.

Environmental monitoring is a complex concept best left to the pros. In this article, we break it down step by step to explain what environmental monitoring evaluates and how these findings are assessed at a professional level.








